2023年12月13日,,華中科技大學同濟醫(yī)學院基礎醫(yī)學院郭崢教授團隊在Cell Press細胞出版社旗下期刊Developmental Cell上以“Eclosion muscles secrete ecdysteroids to initiate asymmetric intestinal stem cell division in Drosophila”為題發(fā)表研究論文。該論文報道了果蠅蛹期的出蛹肌肉DIOM通過分泌蛻皮激素ecdysone起始腸道干細胞不對稱分裂的最新研究成果。基礎醫(yī)學院醫(yī)學遺傳學系博士生張松為論文第一作者。郭崢教授為通訊作者,。

干細胞通過對稱分裂擴增干細胞數(shù)目,,通過不對稱分裂產生終末分化的子代細胞,。在哺乳動物胚胎和器官的發(fā)育過程中,,干細胞總是先進行對稱分裂形成干細胞池,,再通過不對稱分裂產生有功能的子代細胞。例如小鼠腸道發(fā)育中,,胚胎期的腸道干細胞經過數(shù)輪對稱分裂后再進行不對稱分裂,,形成有功能的腸道組織。另外在小鼠大腦發(fā)育過程中,,神經干細胞先通過對稱分裂擴增數(shù)目,,然后轉變?yōu)椴粚ΨQ分裂,產生不同類型的次生神經祖細胞和神經元,。最佳控制論(optimal control theory)認為干細胞的這種先對稱后不對稱的分裂方式可以在最短的時間內形成有功能的組織和器官,。因此,干細胞由對稱分裂到不對稱分裂的轉變時間點是至關重要的,。轉變發(fā)生過早會導致干細胞數(shù)目不足,,難以在有限時間內產生有功能的組織;而轉變時間過晚會導致干細胞的過度增殖和子代細胞的匱乏,。然而,,這個干細胞由對稱分裂轉為不對稱的分子開關是什么?控制這個分子開關的時間在發(fā)育的過程中是如何被決定的,?人們對這些問題都知之甚少,。
在本項研究中,作者利用果蠅蛹期腸道作為研究模型,,探索啟動腸道干細胞由對稱分裂轉向不對稱分裂的信號,。作者利用清華大學果蠅中心的8645株RNAi,通過在pISC中進行全基因組RNAi篩選,,發(fā)現(xiàn)EcR(EcR receptor),、usp(ultraspiracle)和E93(Ecdysone induce protein 93F)這三個蛻皮激素通路 (ecdysone signaling) 基因的敲降能顯著減少出蛹前中腸中EECs的數(shù)量,說明蛻皮激素信號通路起始了pISCs的不對稱分裂,。
作者發(fā)現(xiàn),,EcR與Usp作為蛻皮激素的受體,直接啟動染色體重塑因子E93的轉錄表達,。那么E93是如何起始的pISCs不對稱分裂,?作者把目光集中到ecdysone信號下游的重要靶點Broad (Br) 。Br在蛹初始形成時在腸道表皮有高表達,,但是隨蛹的發(fā)育表達逐漸降低,,在pISCs開始不對稱分裂時Br表達消失(圖1A),這些結果提示沒有Br的表達對于pISCs開啟不對稱分裂至關重要,,而E93關閉了Br的表達,。通過一系列遺傳學實驗,,作者總結出蛻皮激素信號啟動pISC不對稱分裂的分子機制為:EcR/usp通過激活E93關閉Br,使得本被Br抑制的AC-S蛋白復合體和其下游轉錄因子Pros被表達出來,,開啟pISCs的不對稱分裂產生EECs(圖1C),。
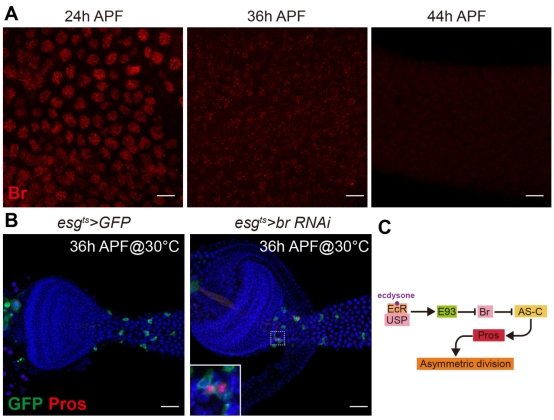
圖1 ecdysone開啟不對分裂的機制
A,44h APF時pISCs開始不對稱分裂,,此時Br不在腸道中表達,。B,在pISCs中敲低Br導致EECs提前產生,。C,,ecdysone信號起始pISCs不對稱分裂的分子機制。
蛻皮激素是如何使得果蠅在44 h APF這個精確的時間點開啟pISC不對稱分裂,? 50年前人們發(fā)現(xiàn)完全變態(tài)昆蟲中蛻皮激素濃度最高峰(脈沖)出現(xiàn)在蛹期的中間時間點,,果蠅的蛻皮激素脈沖出現(xiàn)在40 h APF。作者發(fā)現(xiàn)果蠅pISC在經歷蛻皮激素脈沖后,,隨即開啟不對稱分裂(圖2A),,于是猜測開啟不對稱分裂的時間點是由蛻皮激素在蛹期的脈沖所決定。通過全身性的敲低蛻皮激素合成酶Spookier (Spok) 和Phantom (Phm)后,,蛻皮激素在40 h APF的脈沖濃度顯著降低,,EEC的產生明顯減少(圖2B)。說明果蠅蛹期合成的蛻皮激素脈沖可以起始pISC的不對稱分裂,。
那么蛹期的蛻皮激素脈沖從哪里來的呢,?這是一個困擾研究人員將近百年之久的問題。果蠅在幼蟲期合成蛻皮激素的器官是前胸腺 (PG),。作者發(fā)現(xiàn)在蛹期的PG中敲降蛻皮激素合成酶并不會顯著影響蛻皮激素脈沖的形成,,說明在蛹期除PG外還有關鍵的蛻皮激素來源。作者將T2A-Gal4這個驅動元件通過CRISPR/Cas9技術原位插入到蛻皮激素合成酶phm的編碼序列 3’端后,,發(fā)現(xiàn)Phm除在PG表達外,,還在蛹期的腹部背側內斜肌 (DIOM) 中大量表達,提示脈沖的關鍵組織來源為DIOM (圖2 C),。隨后,,一系列的遺傳和體外培養(yǎng)實驗充分的證明了DIOM在蛹期產生了蛻皮激素的脈沖。
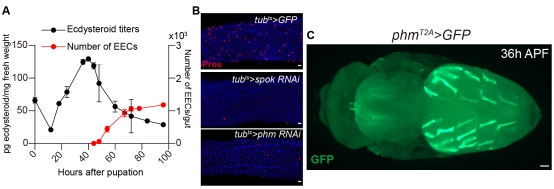
圖2 蛹期ecdysone主要來源于DIOMs
A,,在蛻皮激素高濃度峰后,,腸道開始不對稱分裂產生EECs。B ,,敲低spok和phm兩個蛻皮激素合成酶,,會抑制腸道EECs產生。C,,蛹期phm在PG和DIOMs中表達,。
DIOMs是果蠅在蛹期將幼蟲肌肉重塑成熟后,,用來讓果蠅出蛹的一組肌肉,。作者發(fā)現(xiàn)mTOR信號調控DIOM的重塑,,抑制mTOR后DIOM無法重塑導致蛻皮激素脈沖消失;激活mTOR后DIOM重塑時間提前,,脈沖產生的時間提前,。這些結果表明蛻皮激素脈沖是在DIOM重塑過程中產生。DIOM經過重塑成熟后不再分泌蛻皮激素,,幫助果蠅在羽化時出蛹,,在出蛹后一天內降解。
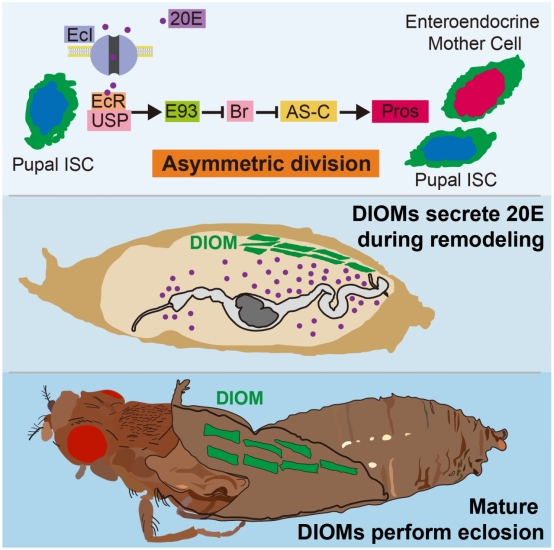
綜上所述,,作者發(fā)現(xiàn)來源于DIOM的果蠅蛻皮激素脈沖將pISC由對稱分裂切換到不對稱分裂,。解決了百年來完全變態(tài)昆蟲在蛹期的蛻皮激素信號來源問題。DIOM作為負責執(zhí)行出蛹的肌肉組織,,其在mTOR信號通路下的重塑成熟過程,,像一個發(fā)育時鐘,通過給予其他器官組織以類固醇激素信號,,精確調控機體各個器官發(fā)育進程,。由于蛻皮激素受體Usp與哺乳動物視黃醇X受體 (RXRs) 同源,作者猜測哺乳動物中視黃酸通路 (retinoid signaling) 調控干細胞由對稱分裂向不對稱分裂的轉變,。
文章鏈接:https://doi.org/10.1016/j.devcel.2023.11.016
 English
English

